


سلام بچه ها
این مقاله خیلی باحال بود، پس تصمیم گرفتم علاوه بر خلاصه ش که اینجا گذاشته بودم، متنشم براتون بزارم. البته اخر مقاله روش تحقیق رو هم اورده بود که کلی فرمول داشت و هر کی خواست قطعا خودش مقاله انگلیسی رو میخونه. ولی مقاله متنش خیلی باحاله. خیلی هم پیچیده نیست و میتونید متنشو بخونید.
حتما اون قسمت که توضیح داده چرا چشم تثبیت بیشتری روی بعضی صحنه ها داره رو جدی بخونید. این جور مقالات برای کسایی که میخوان تبلیغات کنند و یه فیلم تبلیغاتی توی تلوزیون پخش کنند، خیلی کمک کننده است. به دانش پذیرهای تندخوانی نصرت هم کمک میکنه که بفهمن گذراندن زمان بیشتر روی صفحه کتاب، به حفظ شدن بیشتر کمک میکنه یا خیر.
عنوان مقاله: مدت زمان تثبیت در صحنه های طبیعی با رمزگذاری حافظه و نه پردازش تقاضا توضیح داده می شود
نویسندگان: فیلیپ سولوسکی،کارمن امه ،مارتین ان. هبارت ، پیتر کونیگ و تیم سی کیتزمن
سال نشر: 2026
لینک مقاله اصلی:
https://link.springer.com/article/10.1038/s41593-026-02285-1
چکیده
قبل از هر یک از حدود 200000 حرکت چشمی که در روز انجام میدهیم، مغز تصمیم میگیرد قبل از اینکه نگاه خود را به اطلاعات جدید تغییر دهد، چه مدت ثابت بماند. در اینجا ما این فرآیند را با استفاده از یک آزمایش مشاهده صحنه در مقیاس بزرگ (4080 صحنه طبیعی، پنج شرکتکننده) که ترکیبی از مگنتوآنسفالوگرافی، ردیابی چشم و یک کار شرح معنایی است، بررسی میکنیم. با استفاده از تجزیه و تحلیل چند متغیره الگوهای منبع-فضای مگنتوآنسفالوگرافی، تحلیلهای رفتاری و مدلسازی شبکه عصبی مصنوعی (ANN)، نشان میدهیم که تثبیتهای طولانیتر پردازش بصری طولانیمدت را منعکس نمیکنند، بلکه به رمزگذاری حافظه پاییندست مربوط میشوند. اول، تنوع زمانی دینامیک نمایشی جریان شکمی، تنوع در مدت زمان تثبیت را توضیح نمیدهد. دوم، مدت زمان تثبیت با دشواری طبقه بندی پچ تخمین زده شده توسط ANN ضد همبستگی بود. سوم،مدت زمان تثبیت ارتباط مثبتی با به یاد ماندنی پچ پیشبینیشده توسط ANN و گنجاندن عنوان دارد و همزمان با افزایش جفتشدگی فاز-دامنه تتا-گاما، بهویژه در نواحی فرونتال و هیپوکامپ اتفاق میافتد. این نتایج نشان میدهد که تصمیمات زمانبندی حرکت چشم به جای محدودیتهای پردازش ادراکی، بر اساس نیازهای رمزگذاری حافظه شکل میگیرد.
مشاهده محتوای مشابه توسط دیگران

کنترل حافظه کاری مدت زمان تثبیت را در مشاهده صحنه پیش بینی می کند
ماده 26 ژوئیه 2022

یک مدل محاسباتی دو فرآیندی کنترل ثابت-مدت در مشاهده صحنه طبیعی
مقاله دسترسی باز01 سپتامبر 2021

مدت زمان تثبیت در موارد آشنا به دلیل تضعیف کاوش طولانی تر است
مقاله دسترسی باز14 نوامبر 2024
موضوعات مرتبط را کاوش کنید
جدیدترین مقالات، کتاب ها و اخبار در موضوعات مرتبط، پیشنهاد شده با استفاده از یادگیری ماشین را کشف کنید.
- حافظه بلند مدت
- رمزگذاری عصبی
- روان فیزیک
- ساکاد
- ادراک بصری
- حافظه کاری
- توجه بصری و مکانیزم های پردازش شناختی
اصلی
بینایی طبیعی به طور مداوم شامل تصمیم گیری بین حفظ تثبیت فعلی یا تغییر جهت نگاه خود برای کشف اطلاعات بصری جدید است. با توجه به تراکم اطلاعات بالا و تغییرات دینامیکی جهان طبیعی، نمونه برداری سریع از اطلاعات متنوع ارجح به نظر می رسد. با این حال، چشمهای ما در برخی مکانها بیش از 500 میلیثانیه باقی میمانند، در حالی که برخی دیگر برای کمتر از 150 میلیثانیه بازدید میشوند (مراجعههای 1 ، 2 )، که با توجه به سرعت پردازش فوقالعاده سریع مغز، تأخیر قابل توجهی است . این تنوع بزرگ یک سوال اساسی در مورد استراتژی محاسباتی زیربنایی مغز ایجاد می کند.
اگرچه تحقیقات قبلی همبستگی بین مدت زمان تثبیت و ویژگی های تصویر محلی (به عنوان مثال، کنتراست محلی، چگالی لبه)، نیازهای کار و پارامترهای دنباله کاوش 1 ، 2 ، 5 ، 6 ، 7 ، 8 ، 9 را توصیف کرده است ، درک ما از پردازش اطلاعات عصبی زیربنایی محدود است. ما این شکاف را با توسعه پیشبینیکنندههای مرتبط با فرآیند کاهش میدهیم که فرضیههای خاصی را در مورد مکانیسمهای پردازش اطلاعات عصبی که زمانبندی تثبیت را هدایت میکنند، آزمایش میکنند.
یک تئوری رایج در مورد اینکه چرا چشمان ما در برخی مکانها بیشتر استراحت میکنند، مبتنی بر این است که مغز ممکن است به زمانهای متفاوتی برای استخراج اطلاعات نیاز داشته باشد. در واقع، در مورد دید ایستا در ماکاک ها، کار و همکاران. 10 نیاز به پردازش اطلاعات مکرر را در موارد محرکهای چالش برانگیز نشان داد، که نشان میدهد محرکهای بصری پیچیدهتر زمان محاسبات عصبی را طولانیتر میکنند. به طور مشابه، مدلهای شبکه عصبی مصنوعی مکرر (ANN) نشان داده شد که با زمان واکنش انسان در یک کار طبقهبندی انیمیشن 11 مطابقت دارند.. فرضیه پردازش-تقاضای متناظر، مدت زمان تثبیت طولانیتری را برای ویژگیهای تصویر محلی انتظار دارد که شناسایی آنها نسبتاً چالش برانگیز است. در سطح عصبی، این دیدگاه یک همگرایی تاخیری پویایی بازنمایی را هنگام تثبیت اهداف چالش برانگیز در مقایسه با اهداف سادهتر پیشنهاد میکند.
با این حال، یک حساب جایگزین نیز امکان پذیر است. بر اساس این مشاهدات که نمایشهای جریان شکمی فقط یکپارچگی تثبیت متقابل محدود را نشان میدهند ، اطلاعات جریان شکمی تا حد زیادی با حرکت چشمها به یک مکان جدید بازنویسی میشود. بنابراین تثبیت های طولانی مدت می تواند منعکس کننده یک تخصیص زمان استراتژیک مغز برای تثبیت فعال تعبیه عصبی یک پچ ثابت برای پشتیبانی از فرآیندهای پایین دست، مانند رمزگذاری حافظه، قبل از ایجاد اختلال در ساکاد بعدی باشد. ما این گزارش جایگزین از مدت زمان تثبیت را فرضیه تسهیل حافظه می نامیم.
برای تمایز بین این دو فرضیه، یک مجموعه داده مغناطیسی مغزی (MEG) در مقیاس بزرگ جمع آوری کردیم که در آن پنج شرکت کننده به طور فعال 4080 صحنه طبیعی را کاوش کردند (شکل 1 ). این آزمایش شامل یک کار شرح صحنه بود که در طی آن شرکت کنندگان آزادانه هر صحنه طبیعی را به مدت 4 ثانیه کاوش کردند، در 25 درصد موارد و به دنبال آن درخواستی برای ارائه یک توصیف معنایی شفاهی (شکل 1a ). در طول آزمایش، ما به طور همزمان سیگنالهای MEG و حرکات چشم را ضبط میکردیم، که به ما امکان میداد تا پویایی عصبی را که زمان قفل شده برای تثبیتها است، تجزیه و تحلیل کنیم (شکل 1b ). صحنهها از مجموعه دادههای مناظر طبیعی (NSD 13) نمونهبرداری شدند). برای ایجاد یک مجموعه محرک از نظر معنایی متنوع و در عین حال متعادل، جاسازیهای زیرنویس صحنهها را در 60 خوشه معنایی دستهبندی کردیم و به طور یکنواخت از بین آنها نمونهبرداری کردیم (شکل 1c,d ؛ روشها ).
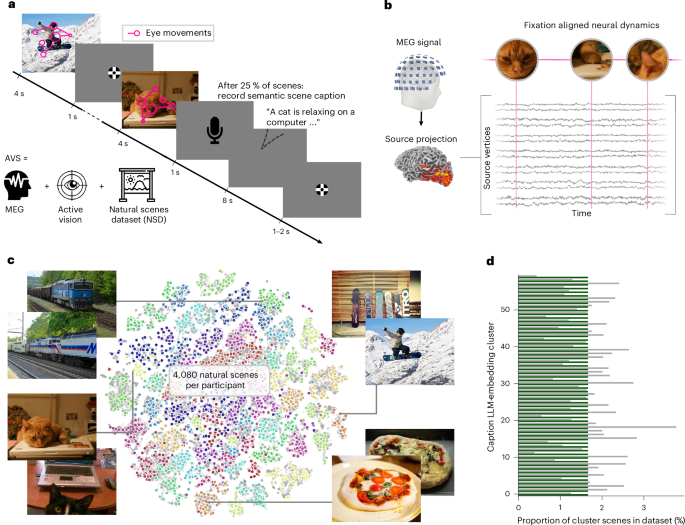
این مجموعه داده در مقیاس بزرگ از MEG و دادههای ردیابی چشم جمعآوریشده در طول دید فعال در صحنههای طبیعی، ما را قادر میسازد تا به طور مؤثر بین این دو فرضیه تمایز قائل شویم. برای آزمایش فرضیه پردازش-تقاضا، پویایی الگوهای فعالسازی منبع MEG را به عنوان تابعی از مدت زمان تثبیت تخمین زدیم و اندازهگیریهای پیچیدگی تصویر را از مدلهای ANN آموزشدیده با کار 11 استخراج کردیم . پردازش پاییندستی مرتبط با حافظه به ازای هر تثبیت بر اساس رتبهبندی صریح اهداف ثابت که در شرح صحنه شرکتکنندگان و امتیازات به یاد ماندنی مبتنی بر ANN 14 ذکر شده است ، استخراج شد که احتمال به خاطر سپردن وصلههای صحنه محلی را تخمین میزند. در نهایت، ما فضای منبع MEG را برای جفت فاز-دامنه تتا-گاما (PAC) 15 ، 16 جستجو کردیم.به عنوان امضای پردازش حافظه
نتایج ما شواهدی را علیه فرضیه پردازش-تقاضا ارائه میکند: دینامیک الگوی عصبی بدون در نظر گرفتن مدت زمان تثبیت در تأخیرهای ثابت تثبیت میشود، و تکههای با طبقهبندی آسانتر تثبیتهای طولانیتری دریافت کردند. در مقابل، اهدافی که بعداً در شرح صحنه ذکر شدند و آنهایی که حافظه پیشبینیشده ANN بالاتری داشتند، تثبیتهای طولانیتری دریافت کردند. تثبیتهای طولانیتر نیز با افزایش تتا-گاما PAC در نواحی فرونتال و هیپوکامپ اتفاق افتاد. این یافتهها با هم نشان میدهند که زمانبندی تثبیت، تخصیص زمان استراتژیک برای رمزگذاری حافظه را به جای نیازهای پردازش بصری منعکس میکند.
نتایج
تأخیر ثابت تثبیت الگوی MEG بدون در نظر گرفتن مدت زمان تثبیت
ابتدا، بررسی کردیم که آیا دینامیک زمانی متفاوت الگوهای فعالیت عصبی میتواند تفاوتها در مدت زمان تثبیت را توضیح دهد (شکل 2a,b ). برای هر ناحیه بصری مورد علاقه (ROI)، فاصله همبستگی بین بردارهای فعال سازی متوالی را با یک شکاف زمانی متحرک 30 میلی ثانیه محاسبه کردیم. این اندازه گیری چگونگی تغییر الگوهای فعالیت عصبی را در طول زمان تعیین کرد (شکل 2c ). نکته قابل توجه، نقطه تثبیت الگوی عصبی – که بهعنوان نقطه نیمه راه پهلوی در حال سقوط به دنبال موج اصلی تغییر الگو مشخص میشود – در مقاطع زمانی مشابه، بدون در نظر گرفتن مدت زمان تثبیت بعدی (شکل 2d ) برای همه مناطق بینایی آزمایششده، از جمله نواحی اولیه بینایی، شکمی، جانبی و جداری رخ داده است. (شکل 2e). یک مدل خطی با اثرات مختلط با وقفههای تصادفی برای شرکتکنندگان و کنترل نیمکره و ROI نشان داد که زمانبندی تثبیت الگو تا حد زیادی مستقل از مدت زمان تثبیت بود (همه P > 0.25).
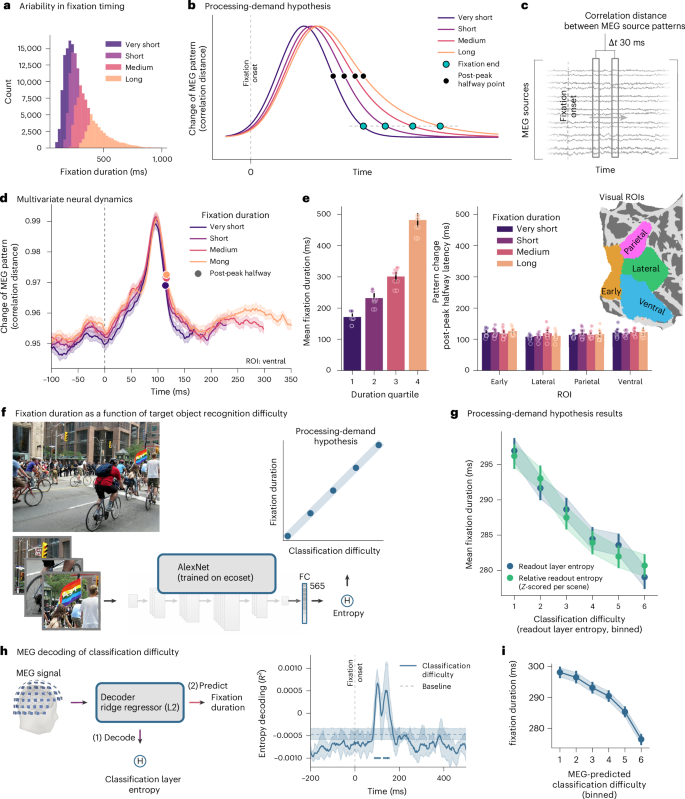
مشاهده اینکه میانگین تأخیر نقاط نیمه راه در هر چارک مدت با میانگین مدت تثبیت در هر چارک مقیاس نمیشود، نشان میدهد که بازنماییهای عصبی در زمانبندی ثابت به حالت نسبتاً پایداری میرسند، مستقل از مدت زمانی که تثبیت طول میکشد. این یافته در تضاد با فرضیه پردازش-تقاضا است، که تثبیت تاخیری را برای تثبیتهای طولانیتر که نیاز به پردازش مکرر طولانیتری دارند، پیشبینی میکند.
اهداف ساده تر به جای چالش برانگیز، تثبیت طولانی تری دریافت می کنند
برای بررسی بیشتر فرضیه پردازش-تقاضا، ما تجزیه و تحلیل کردیم که چگونه دشواری طبقهبندی وصلههای تصویر ثابت شده با مدت زمان تثبیت ارتباط دارد. برای تخمین دشواری طبقهبندی، آنتروپی لایه طبقهبندی یک AlexNet مدل 17 آموزشدیده بر روی ecoset 18 را محاسبه کردیم ، که در آن آنتروپی کمتر نشاندهنده اطمینان طبقهبندی بالاتر 11 است (شکل 2f).). برای اطمینان از عدم توزیع محصولات مبتنی بر تثبیت برای ANN آموزشدیدهشده، مدل را بر روی یک نسخه اصلاحشده از ecoset آموزش دادیم، با تمرکز بر بخشهای تصویر مطابق با اندازه حاوی دستههای ecoset مربوطه. در مقایسه آنتروپی طبقه بندی با مدت زمان تثبیت، ما یک رابطه منفی پیدا کردیم، که نشان می دهد وصله های طبقه بندی آسان تر طولانی تر ثابت می شوند (شکل 2g ، خط آبی). مدلهای خطی با جلوههای مختلط، بر روی مقادیر آنتروپی با log تبدیل شده ، تأیید کردند که آنتروپی طبقهبندی بالاتر تثبیتهای کوتاهتری را پیشبینی میکند ( β = -5.44 ms، 95٪ فاصله اطمینان (CI): (6.07-، -4.81)، P <0.001؛ n = 162020 تثبیت، پنج شرکت کننده). نکته مهم این است که این اثر توسط تفاوتهای سطح صحنه ایجاد نمیشود، زیرا این رابطه در هنگام محاسبه آنتروپی طبقهبندی نسبی ( با امتیاز Z ) در تمام وصلههای بازدید شده در یک صحنه مشخص ادامه داشت ( β = -5.13 میلیثانیه، 95٪ فاصله اطمینان (CI): (-5.76، -4.50)، P <0.02g، خط سبز). این تجزیه و تحلیل اخیر تأیید کرد که اثرات مشاهده شده در حالی که در سطح تثبیتهای فردی عمل میکنند، به زمینه حساس هستند.
برای ارائه اعتبار عصبی رابطه بین دشواری تشخیص و مدت زمان تثبیت، از سیگنالهای MEG برای رمزگشایی آنتروپی طبقهبندی مشتق شده از AlexNet استفاده کردیم. ما رمزگشاهای رگرسیون برجستگی را بر روی فعالیت MEG هم تراز با تثبیت آموزش دادیم تا مقادیر آنتروپی لایه طبقه بندی تکه های ثابت را رمزگشایی کنیم (شکل 2h ). دقت رمزگشایی در 114.4 ms (95% CI: (98.0، 130.8)) پس از تثبیت ( R2 = 0.001، 95% CI: (0.001، 0.002)، P <0.001، P <0.001) به اوج رسید. به طور اساسی، هنگام استفاده از این رمزگشا برای پیشبینی آنتروپی طبقهبندی از توپوگرافیهای گرادیومتر، آنتروپی پیشبینیشده با MEG بالاتر را برای پیشبینی تثبیتهای کوتاهتر مشاهده کردیم ( β = -3.89 ms، 95٪ فاصله اطمینان (CI): (-4.68، -3.10)، P <0.001)، مطابق با مشاهدات رفتاری قبلی ما. رابطه منفی بین دشواری طبقه بندی و مدت زمان تثبیت در چندین معماری شبکه عصبی کانولوشنال تعمیم یافته است (AlexNet، VGG16، Inception-v3، ResNet50؛ شکل تکمیلی 1 )، که نشان دهنده استحکام این اثر است. در مجموع، بر خلاف فرضیه پردازش-تقاضا، که تثبیتهای طولانیتر را برای ورودیهای بصری چالشبرانگیز پیشبینی میکند، نتایج ما الگوی مخالف را نشان داد: مدت زمان تثبیت برای وصلههای تصویری که تشخیص آسان به نظر میرسید طولانیتر بود.
تثبیت های طولانی تر، استفاده پایین دستی از اطلاعات بصری را پیش بینی می کند
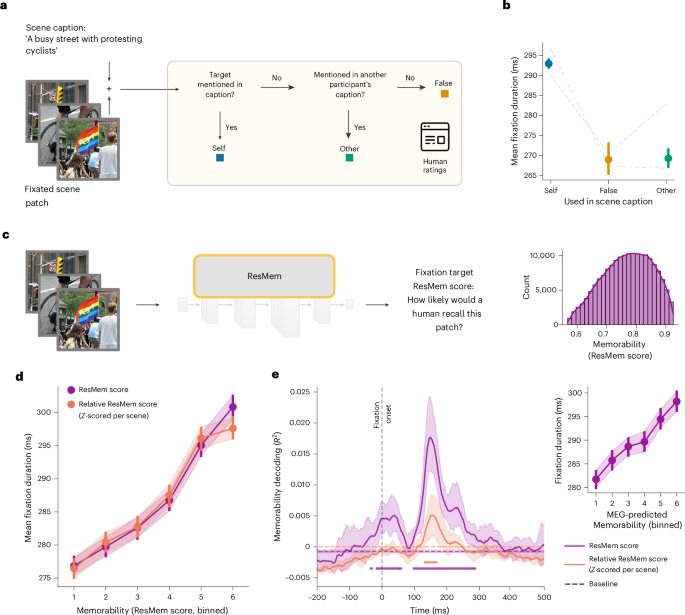
برای آزمایش فرضیه تسهیل حافظه، ما پرسیدیم که آیا تثبیت بر روی اشیایی که بعداً به عنوان بخشی از شرح صحنه شرکت کنندگان گزارش شد، در مقایسه با اشیای مشاهده شده اما گزارش نشده، طولانیتر ثابت میشوند یا خیر. برای هر هدف تثبیت، دو ارزیاب مستقل ارزیابی کردند که آیا بعداً در توصیف صحنه شرکتکننده به آن اشاره شده است یا خیر. برای اهدافی که توسط شرکتکننده ذکر نشده است، ارزیابیکنندگان بهعلاوه ارزیابی کردند که آیا سایر شرکتکنندگان آنها را در زیرنویسهای خود گنجاندهاند یا خیر (شکل 3a ). ما متوجه شدیم که میانگین مدت تثبیت برای اهداف تثبیت که بعداً در عنوان خود شرکتکننده به آنها اشاره شد (293.0 ms، 95٪ فاصله اطمینان (CI): (292.0، 294.0)) در مقایسه با موارد ذکر نشده (269.0 ms، 95٪ فاصله اطمینان: (265.2، 265.2، 272.9٪، 272.9٪، هدف هرگز ذکر نشده) بیشتر است. CI: (267.2، 271.4) برای اهدافی که فقط توسط دیگران ذکر شده است.3b ). یک مدل با اثرات مختلط ( n = 86738 تثبیت) نشان داد که اهداف خودارجاعی به طور قابلتوجهی تثبیت طولانیتری نسبت به هر دو هدف غیر مرجع داشتند ( β = 25.7 ms، 95% فاصله اطمینان (CI: (21.3، 30.1)، P <0.001) و اهدافی که فقط توسط سایر شرکتکنندگان ذکر شده است. 23.7 )، P <0.001). به طور بحرانی، میانگین مدت سرمایه گذاری روی اهدافی که فقط توسط دیگران ذکر شده است تفاوت قابل توجهی با اهداف غیر مرجع نداشت ( β = 4.7 ms، 95٪ فاصله اطمینان (CI): (0.3-، 9.6)، 0.064 = P )، که نشان می دهد این اثر به استفاده بعدی شرکت کنندگان از اطلاعات بصری به جای ویژگی های هدف بصری اختصاص دارد.
اهداف تثبیت به یاد ماندنی مدت زمان تثبیت طولانی تری دریافت می کنند
برای گسترش تحلیلهای خود فراتر از تثبیتها روی ۲۵٪ صحنههایی که با کار شرحنویسی برای همه تثبیتهای ضبطشده دنبال میشد، به خاطر سپردن پچ را با استفاده از یک ANN آموزشدیده برای پیشبینی حافظه انسانی ۱۴ از مجموعه دادههای حافظه بصری رفتاری در مقیاس بزرگ تخمین زدیم (LaMem 19 و MemCat 20 ؛ شکل 3c ). این مدل عملکرد حافظه بصری را که در بازههای زمانی بین 27 ثانیه تا 3.5 دقیقه پس از 35 تا 150 تصویر مداخلهای آزمایش شده است، ثبت میکند. بنابراین، تخمینهای به یاد ماندنی اجازه میدهند تا زمان تثبیت مشاهدهشده را با تخمینهای پردازش حافظه برای مقیاسهای زمانی مرتبط کنیم که از جفت شدن مستقیم زمانی کاوش صحنه و کار زیرنویس بعدی فراتر میرود.
مطابق با نتایج قبلی ما، میانگین مدت تثبیت را بیش از شش سطل (جنسی) افزایش دادیم که تخمینهای به یاد ماندنی فزاینده را به تصویر میکشد (شکل 3d ، خط سرخابی). این رابطه برای تخمینهای خاطرهانگیز بودن نسبی ادامه داشت: یعنی پس از عادیسازی تخمینهای خاطرهانگیز بودن بر روی مجموعه تمام اهداف بازدید شده در یک صحنه (شکل 3d ، خط نارنجی). دومی نشان میدهد که اثرات مشاهدهشده در سطح تثبیت خاص (حساس به زمینه) عمل میکنند، بهجای اینکه منعکسکننده به یاد ماندنیبودن کلی صحنه باشند. مدلهای خطی با جلوههای مختلط با وقفههای تصادفی برای شرکتکنندگان تأیید کردند که هم حافظهپذیری مطلق ( β = 8.05 میلیثانیه، 95٪ فاصله اطمینان (CI): (7.43، 8.66)، P <0.001) و هم خاطرهانگیزی نسبی ( β). = 7.70 میلیثانیه، 95% فاصله اطمینان (CI): (7.08، 8.31)، P <0.001) تثبیتهای طولانیتری را پیشبینی کردند ( n = 168946 تثبیت، پنج شرکتکننده). رابطه مثبت بین به یاد ماندنی و مدت تثبیت در بین مقوله های معنایی قوی بود، با اثرات قابل توجهی برای اهداف تثبیت جاندار و بی جان (همه P <0.001؛ شکل تکمیلی 2 ). روی هم رفته، اهدافی که پیش بینی می شد به یاد ماندنی تر باشند، تثبیت های طولانی تری دریافت کردند.
کار قبلی نشان داده است که مدت زمان تثبیت نیز زمانی افزایش مییابد که ساکاد بعدی مکان صحنه دورتر را هدف قرار دهد . هنگامی که دامنه ساکاد را در مدل ما لحاظ کنیم، اثر به یاد ماندنی قوی باقی ماند ( β = 8.07 میلیثانیه، 95% فاصله اطمینان (CI) (7.45، 8.69)، Z = 25.50، P <0.001)، در حالی که دامنه ساکاد همچنین تثبیتهای طولانیتری را پیشبینی میکند ( β = 2.7٪، Z ، 39/2، 39 = ms) 7.54، P <0.001). قابل ذکر است، ما یک تعامل قابل توجه بین به یاد ماندنی و دامنه ساکاد مشاهده کردیم ( β = 2.74 ms، 95٪ CI (2.12، 3.36)، Z = 8.66، P <0.001)، نشان می دهد که اثر به یاد ماندنی برای تثبیت های قبل از ساکادهای بزرگتر قوی تر بود. این الگو نشان می دهد که رمزگذاری حافظه به طور استراتژیک توسط برنامه های موتوری آینده تعدیل می شود و از یک فرآیند رمزگذاری فعال به جای گلوگاه های پردازش غیرفعال پشتیبانی می کند.
تجزیه و تحلیل رمزگشایی از خاطرهانگیز بودن، که در طول زمان تخمین زده شد، نشان داد که خاطرهانگیز بودن میتواند با موفقیت از الگوهای فعالسازی گرادیومتر خوانده شود. برای به خاطر سپردن مطلق، اوج دقت رمزگشایی ( R2 = 0.020، 95% CI: (0.015، 0.026)) در 148.8 ms (95% CI: (144.0، 154.0)) پس از تثبیت رخ داد. برای به خاطر سپردن نسبی، حداکثر دقت رمزگشایی ( R2 = 0.007، 95% فاصله اطمینان (CI ) : (0.003، 0.010)) در 122.0 ms (95% فاصله اطمینان: (57.2، 158.4)) پس از تثبیت-شروع ( N = 3 شرکت کننده؛) رخ داد .). به طور بحرانی، برای ارائه اعتبار عصبی رابطه بین به خاطر سپردن و مدت زمان تثبیت، از خروجی رمزگشای حافظه عصبی استفاده کردیم و آن را به مدت زمان تثبیت مرتبط کردیم. ما خاطرهانگیزی پیشبینیشده با MEG را برای تثبیتهای طولانیتر مشاهده کردیم ( β = 5.62 میلیثانیه، 95٪ فاصله اطمینان (CI): (5.01، 6.23)، P <0.001). این نتایج نشان میدهد که اطلاعات به یاد ماندنی به زمانبندی تثبیت و الگوهای فعالیت عصبی مربوط میشود، که نشان میدهد فرآیندهای حافظه برای زمانبندی تثبیت بینایی فعال مرتبط هستند.
جفت شدن تتا-گاما در دینامیک عصبی در طول تثبیت طولانی تر
به دنبال رویکرد داده محور ما که رابطه بین مدت زمان تثبیت و به خاطر سپردن را شناسایی کرد، ما یک تجزیه و تحلیل هدفمند انجام دادیم که در آن داده های MEG هم تراز با تثبیت را برای PAC بین نوسانات تتا و گاما، یک امضای عصبی از پردازش مرتبط با حافظه ، تجزیه و تحلیل کردیم . ما تحلیلهای اولیه خود را بر روی تثبیتهای طولانیتر با حداقل 300 میلیثانیه متمرکز کردیم و PAC را در یک پنجره در طول 250 میلیثانیه نهایی قبل از افست تثبیت محاسبه کردیم (شکل 4a ). این رویکرد آفست قفل شده، فعالیت عصبی را قبل از تصمیم برای پایان دادن به تثبیت ثبت می کند.
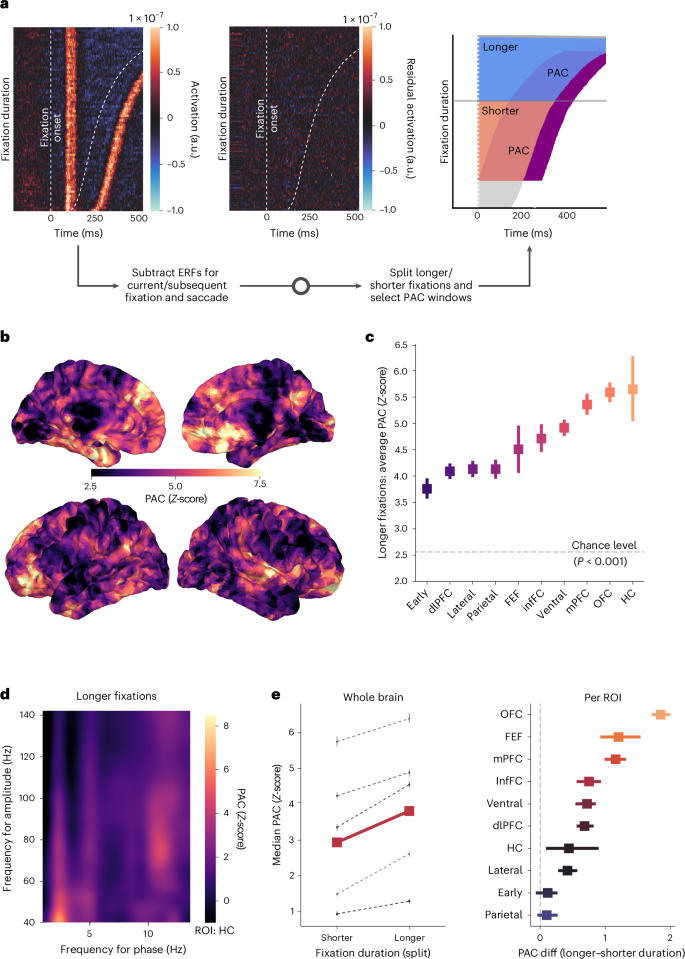
برای جداسازی کوپلینگ نوسانی از پاسخهای برانگیخته، میدانهای مرتبط با رویداد (ERFs) ساکاد فعلی و تثبیت و همچنین ساکاد و تثبیت بعدی (باقیماندههای چهار برابر؛ شکل 4a ) را کم کردیم. برای اطمینان از استحکام آماری و کنترل جفت جعلی، ما از روش جانشین تک برشی 15 استفاده کردیم که ساختار زمانی دادهها را حفظ میکند و در عین حال روابط فاز- دامنه خاص را مختل میکند.
در تضاد با اندازهگیریهای PAC مشاهدهشده ما در برابر این توزیع تهی جایگزین، جفت شدن تتا-گاما را در چندین ناحیه قشر مغز شناسایی کردیم (شکل 4b ). در مجموعه گسترده ای از ROI، ما PAC قابل توجهی را در منابع بیشتر به ازای هر ROI نسبت به آنچه به صورت تصادفی انتظار می رفت مشاهده کردیم ( تست های t یک نمونه ، تصحیح شده با FDR، همه P <0.05)، که جفت شدن تتا-گاما را بالاتر از سطوح شانس در سراسر سلسله مراتب قشر مغز نشان می دهد.
یک مدل اثرات مختلط که امتیازات PAC Z را در نواحی مغزی مقایسه میکند (با شرکتکننده بهعنوان اثر تصادفی) جفتشدگی قابلتوجهی قویتری را در مقایسه با قشر بینایی اولیه نشان داد (همه β ≥ 0.23، همه P <0.05) در قشر جانبی و جداری، میدان چشم پیشانی، قشر میانی پیشانی جلویی، جلوی قشر پیشانی، قشر بینایی تحتانی. نواحی قشر (mPFC)، قشر جلوی پیشانی پشتی جانبی (dlPFC)، قشر اوربیتوفرونتال (OFC) و هیپوکامپ (HC) (شکل 4c ). برای تأیید استحکام یافتههای PAC در پنجره زمانی مورد استفاده برای تجزیه و تحلیل، ما این تحلیل را با یک پنجره قفلشده افست طولانی (نهایی 350 میلیثانیه قبل از پایان تثبیت، با تقسیم مدت زمان در 350 میلیثانیه؛ شکل تکمیلی 4) تکرار کردیم.). تجزیه و تحلیل PAC فرکانس به فرکانس جداگانه یک پنجره گسترده و وضوح بهتری از باندهای درگیر در PAC مشاهده شده ارائه می دهد (شکل 4d ).
برای آزمایش مستقیم اینکه آیا چنین الگوهای کوپلینگ مرتبط با حافظه در طول تثبیتهای طولانیتر افزایش مییابند، ما قدرت PAC را بین تثبیتهای طولانیتر (> 300 میلیثانیه) و کوتاهتر (کمتر از 300 میلیثانیه) مقایسه کردیم، با هر دو شرایط با استفاده از پنجره قفلشده با افست یکسان (250 میلیثانیه پایانی قبل از آفست تثبیت؛ شکل 4a ). این مقایسه جفت شدن تتا-گاما قویتر را در مدت طولانیتر در مقایسه با تثبیتهای کوتاهتر نشان داد (شکل 4e ). تفاوتهای مدت زمان آزمایش مدل با اثرات مختلط (طولانیتر – کوتاهتر) در هر ROI نشاندهنده PAC قویتر در طول تثبیتهای طولانیتر در قشر فرونتال است: OFC ( β = 1.99، 95٪ CI (1.39، 2.60)، P <0.001)، میدان چشم فرونتال ( β = 1.14، 1.3، 09.5 = 1.14، 1.5، 9. 0.002 )، mPFC (β = 0.97، 95% CI (0.37، 1.58)، P = 0.002)، infFC ( β = 0.95، 95% CI (0.34، 1.57)، P = 0.002) و dlPFC ( β = 0.74، 93% CI، 93، 0.6 = 0.7) همچنین در قشر شکمی ( β = 0.69، 95٪ CI (0.09، 1.29)، P = 0.024). با هم، این نتایج نشان میدهد که تثبیتهای طولانیتر با امضاهای عصبی تقویتشده رمزگذاری و نگهداری حافظه همزمان اتفاق میافتد و شواهد عصبی مستقیمی را برای پردازش مرتبط با حافظه در طول مشاهده طولانی ارائه میدهد.
بحث
با استفاده از مجموعه دادههای MEG در مقیاس بزرگ در ارتباط با ردیابی چشم، تحلیلهای رفتاری و مدلسازی محاسباتی، نشان میدهیم که مدت زمان تثبیت طولانیتر با پردازش پاییندستی مرتبط با حافظه به جای نیازهای پردازش بصری بهتر توضیح داده میشود. پویایی الگوی عصبی بدون در نظر گرفتن مدت زمان تثبیت نهایی در مقاطع زمانی مشابه تثبیت شد و شرکتکنندگان به جای وصلههای تصویر پیچیدهتر، برای طبقهبندی آسانتر به مدت طولانیتری تثبیت کردند. این رابطه منفی بین دشواری طبقهبندی و مدت زمان تثبیت در دادههای عصبی هنگام استفاده از یک مدل رمزگشایی برای پیشبینی دشواری طبقهبندی مستقیماً از سیگنال MEG حفظ و تأیید شد.
این مجموعه از یافتهها در تضاد با شواهد قبلی از آزمایشهای مشاهده استاتیک ماکاک است که در آن محرکهای چالشبرانگیز به پردازش مکرر طولانیمدت نیاز دارند (به عنوان مثال، رجوع به 10 ). در عوض، دادههای ما نشان میدهند که یک رابطه پیچیدهتر در طول دید فعال در صحنههای طبیعی، جایی که فرآیندهای پاییندستی بیشتر، مانند عوامل مرتبط با حافظه، تأثیر قابلتوجهی بر مدت زمان تثبیت دارند.
چهار یافته کلیدی از دخالت فرآیندهای مرتبط با حافظه به عنوان محرک طولانی مدت تثبیت پشتیبانی می کنند: (1) اشیایی که بعداً در شرح صحنه ذکر شدند، تثبیت های طولانی تری دریافت کردند. (2) اهدافی که پیشبینی میشد از طریق یک شبکه عمیق آموزشدیده بر روی دادههای انسانی، قابلیت به خاطر سپردن بالاتری داشته باشند، به طور مداوم تثبیتهای طولانیتری دریافت کردند. (3) خاطرهانگیز بودن (نسبی) تکههای ثابت با موفقیت از دادههای MEG رمزگشایی شد، با به خاطر سپردن رمزگشایی بالاتر که تثبیتهای طولانیتری را پیشبینی میکند. و (4) افزایش تتا-گاما PAC در طول تثبیت طولانی تر مشاهده شد.
این برهمکنش نوسانی، که در آن دامنه گاما توسط فاز تتا مدوله میشود، به طور قوی به حافظه کاری (بصری) رمزگذاری 21 ، 22 ، 23 ، 24 ، 25 ، 26 ، 27 ، 28 ، 29 مرتبط شده است . در این کار قبلی، جفت شدن تتا-گاما عمدتاً در قشر پیشانی، HC و نواحی جداری مشاهده شد، که با الگوی توپوگرافی جفت تتا-گاما که مشاهده کردیم، همسو بود.
اگرچه PAC در زمینههایی مشاهده شده است که به فرآیندهای حافظه محدود نمیشود 26 ، 30 ، چندین مشاهدات در اینجا از یک تفسیر یادگاری پشتیبانی میکنند. (1) توزیع فضایی PAC تقویتشده در طول تثبیتهای طولانیتر، قویترین در نواحی فرونتال و هیپوکامپ به جای قشر بینایی، با شبکههای حافظه همسو میشود 23 ، 31 ، 32. به طور بحرانی، (2) تثبیتهای طولانیتر هم با خاطرهانگیزی بالاتر و هم با گزارشهای بعدی در توصیف صحنه همراه است. مدولاسیون سیستماتیک PAC در طول مدت زمان تثبیت، همراه با این پیوند رفتاری با نتایج حافظه، از تفسیر مرتبط با حافظه در زمینه فعلی پشتیبانی میکند. تجزیه و تحلیل PAC حاضر با استفاده از داده های MEG تراز شده با تثبیت در طول کاوش صحنه طبیعی، در قلمروی عمدتا ناشناخته عمل می کند. برای مثال، ERF های مرتبط با شروع تثبیت، تغییرات طیفی گذرا را ایجاد می کنند که ممکن است بر تخمین PAC تأثیر بگذارد. روش تفریق چهارگانه ERF، آزمایش جایگزین و تضاد مدت زمان بین تثبیتهای طولانیتر در مقابل کوتاهتر، بنابراین کنترلهای مهمی را فراهم میکند. با این حال، کار آینده نیاز به بررسی بیشتر نشانههای عصبی دقیق حافظه فعال در طول بینایی فعال دارد.
یافتههای ما از اطلاعات بصری نسبتاً تثبیتشده با مشاهدات شیائو و همکارانش همسو میشود و آن را گسترش میدهد. 12او دریافت که جمعیت های عصبی در جریان شکمی هر تثبیت را به طور مستقل پردازش می کنند و هیچ یکپارچه سازی اطلاعات را در سراسر ساکاد مشاهده نمی کنند. چنین پردازشهای خاص تثبیت در جریان شکمی ممکن است نیاز به تثبیتهای طولانیتر در مناطق تصویر را توضیح دهد که حاوی اطلاعات مورد نیاز برای فرآیندهای پاییندست بعدی است، زیرا تثبیت در جای دیگر سیستم را با اطلاعات جدید “شباع” میکند. در واقع، دادههای ما نشان میدهد که جفت شدن تتا-گاما عمدتاً در نواحی جلویی قدامی، اوربیتوفرونتال و هیپوکامپ رخ میدهد تا در نواحی بینایی. این تفکیک فضایی یک تقسیم کار را پیشنهاد میکند: نواحی جریان شکمی بصری تثبیتهای فعلی را پردازش میکنند، در حالی که مناطق درجه بالاتر، در صورت نیاز، اطلاعات را در سراسر حرکات چشم یکپارچه میکنند .، امکان کاوش بصری سریع را فراهم می کند و در عین حال از درک یکپارچه صحنه بصری کلی و زمینه سازی اطلاعات محلی پشتیبانی می کند.
هنگام کاوش در محیط بصری، ساکادهای بزرگتر مستلزم جابجایی فضایی بیشتر است که به طور بالقوه تجمیع اطلاعات بصری را قبل از خروج از مکان فعلی مهم تر می کند. این یافته که محتوای به یاد ماندنی باعث تثبیت نسبتا طولانیتر قبل از ساکادهای بزرگتر میشود، نشان میدهد که رمزگذاری به طور استراتژیک توسط برنامههای حرکتی آینده تعدیل میشود. این الگو با یک فرآیند رمزگذاری فعال و انعطاف پذیر به جای گلوگاه های پردازش غیرفعال سازگار است، جایی که سیستم به طور تطبیقی زمان رمزگذاری را بر اساس ارزش محتوا و جابجایی آتی تخصیص می دهد.
این دیدگاه کار اخیر را گسترش می دهد که نشان می دهد یک تعامل هماهنگ بین حرکات چشم و فرآیندهای حافظه است. مطالعات قبلی نشان دادهاند که اجرای ساکاد میتواند بر حفظ حافظه کاری تأثیر منفی بگذارد 34 ، 35، در حالی که با الگوهای هیپوکامپ که شکلگیری حافظه را تقویت میکنند، هماهنگ میشود . بعلاوه، افرادی که ظرفیت حافظه کاری بالاتری دارند، مدت زمان تثبیت طولانی تری را در حین مشاهده صحنه نشان می دهند 38 . این هماهنگی بین حرکات چشم و فرآیندهای حافظه با چارچوبهای کدگذاری پیشبینیکننده 39 سازگار است ، که در آن حرکات چشم را میتوان به عنوان آزمایشهایی توصیف کرد که فرضیههای مربوط به محتوای بصری را آزمایش میکنند .. در اینجا نمایشهای حافظه ، پیشبینیهایی را درباره اطلاعات بصری در مکانهای مختلف راهنمایی میکنند، که نمونهبرداری کارآمد از مناطق اطلاعاتی و بهروزرسانی مدلهای صحنه داخلی را امکانپذیر میسازد .
اگرچه تجزیه و تحلیل ما بینش های مکانیکی را در مورد فرآیند تصمیم گیری نشان می دهد که چگونه مغز تصمیم می گیرد چه زمانی اطلاعات را از یک مکان جدید نمونه برداری کند، تغییر کلی مشاهده شده در مدت زمان تثبیت تا حدی غیرقابل توضیح است که احتمالاً منعکس کننده مخلوطی از فرآیندهای قطعی و تأثیرات تصادفی است 2 ، 44 ، 45 . کار آینده با استفاده از مدلهایی که درک معنایی 46 یا ویژگیهای مربوط به عمل را در بر میگیرد ، میتواند بینشهای بیشتری را در مورد اینکه چگونه مغز تخصیص تثبیت را در طول دید طبیعی تعیین میکند، ارائه دهد.
مجموعه داده ما بر روی پنج شرکتکننده متمرکز است ، اما این طراحی فشرده درون موضوعی از رویکرد NSD 13 و سایر مجموعههای داده پیشرفته پیروی میکند که قابلیت اطمینان و قدرت آماری را از طریق نمونهگیری گسترده از مغزهای فردی در اولویت قرار میدهند . فرآیندهای ادراکی و حافظه که در اینجا مورد بررسی قرار میگیرند احتمالاً مکانیسمهای اساسی را منعکس میکنند که در بین افراد تعمیم مییابد، همانطور که توسط اثرات ثابت مشاهده شده در همه شرکتکنندگان پیشنهاد میشود. با این وجود، مطالعات آینده با گروههای بزرگتر و متنوعتر از شرکتکنندگان برای تعمیم این یافتهها مفید خواهد بود.
یافتههای ما نشان میدهد که تنوع قابلتوجه در زمانبندی تثبیت در درجه اول نه به دلیل پیچیدگی پردازش، بلکه بیشتر به دلیل الزامات پردازش پایین دست است. نمایشهای بصری در مقاطع زمانی ثابت بدون توجه به مدت زمان تثبیت تثبیت میشوند و با حسابهای پردازش-تقاضا متناقض هستند. در مقابل، جفت شدن تتا-گامای مرتبط با حافظه در مدت طولانیتر در مقایسه با تثبیتهای کوتاهتر قویتر بود. این نتایج بینایی فعال را بهعنوان حافظه محور ایجاد میکنند، جایی که تثبیتهای طولانیتر منعکسکننده سرمایهگذاری زمانی استراتژیک برای تثبیت کدهای عصبی قبل از اختلال ساکادیک است. بنابراین مغز مدت زمان نمونه برداری را بر اساس ارزش اطلاعات، نه پیچیدگی محاسباتی، بهینه می کند.
تحلیلهای ما از مجموعه دادههای MEG و ردیابی چشم در مقیاس بزرگ نشان میدهد که مورد بینایی فعال نیاز به یک تغییر پارادایم در علوم اعصاب بصری دارد. در چشم انداز فعال، انسان ها به طور استراتژیک منابع زمانی را بر اساس مطلوبیت پایین دستی به جای نیازهای پردازش فوری تخصیص می دهند. این یافته راه های جدیدی را برای درک جمع آوری اطلاعات تطبیقی با مفاهیمی که از علم بینایی به سایر حوزه های حسی و شناختی گسترش می یابد، باز می کند. بهطور گستردهتر، این بینشها درک ما را از نحوه مدیریت سیستمهای هوشمند مبادلات اکتشاف و بهرهبرداری نشان میدهد.

بیسبال ممکن است سرگرمی مورد علاقه آمریکا باشد، اما در رتبه بندی محبوب ترین سرگرمی ها، خواندن به طور مداوم رتبه بالایی دارد . فهمیدن دلیل آن سخت نیست: خواندن همزمان جذاب و آرامش بخش است و انجام آن به تنهایی و با دوستان لذت بخش است.
تیمی از محققان در موسسه علوم و فناوری پیشرفته بکمن دلیل دیگری را برای علاقه به مطالعه کشف کرده اند: ممکن است به حفظ مهارت های حافظه با افزایش سن افراد – و مغز آنها – کمک کند. کار آنها در Frontiers in Psychology گزارش شده است .
لیز استاین-مورو ، محقق بکمن ، که همچنین مدیر آزمایشگاه آموزش بزرگسالان است ، گفت: «مطالعه اوقات فراغت، نوعی که واقعاً شما را جذب میکند، برای شما مفید است و به ایجاد تواناییهای ذهنی که خواندن به آن بستگی دارد کمک میکند. عضو هیئت علمی گروه روانشناسی تربیتی و محقق ارشد مطالعه.
یکی از این تواناییهای ذهنی، حافظه اپیزودیک یا حافظه برای رویدادها است که به ما امکان میدهد آنچه را در فصلهای قبلی یک کتاب اتفاق افتاده به خاطر بیاوریم و داستان در حال انجام را درک کنیم. توانایی دیگر حافظه فعال است، ظرفیت نگه داشتن چیزها در ذهنمان در حالی که درگیر فرآیندهای ذهنی دیگر هستیم. حافظه فعال به ما کمک می کند تا در ادامه خواندن، چیزهایی را که در پاراگراف های اخیر رخ داده است، پیگیری کنیم.
هم حافظه اپیزودیک و هم حافظه کاری با افزایش سن کاهش می یابد، اما خوانندگان معمولی به طور معمول این مهارت ها را در زمینه های مختلف تمرین می کنند.
ادبیات بسیار قوی وجود دارد که نشان می دهد بین حافظه فعال و درک زبان و حافظه بلند مدت رابطه وجود دارد. به نظر می رسد حافظه کاری با افزایش سن کاهش می یابد، اما تغییرات زیادی به خصوص در میان افراد مسن وجود دارد.
رمز و راز پیرامون رابطه بین خواندن و حافظه این است که آیا خواندن به بهبود حافظه کمک می کند یا اینکه توانایی های حافظه کاری قوی مهارت های درک مطلب را بهبود می بخشد. دانستن جهت علیت پیامدهای مهمی بر انواع درمان هایی خواهد داشت که می تواند به حفظ حافظه در طول زندگی کمک کند.
استاین-مورو و تیم بین رشته ای که شامل محققان بکمن، دکتر دانیل لانو ، استاد فیزیولوژی مولکولی و یکپارچه، و آرون باربی ، استاد روانشناسی بود، مطالعه ای را برای آزمایش رابطه علی بین خواندن و حافظه انجام دادند. برای شروع، آنها به مجموعهای از کتابهای جالب و جذاب نیاز داشتند – از آن دستههایی که واقعاً شما را میکشد – و تصمیم گرفتند با کارشناسان بخش خدمات بزرگسالان کتابخانه عمومی Champaign تماس بگیرند.

کریستینا هورنر، مدیر خدمات بزرگسالان کتابخانه در زمان مطالعه گفت: «ما صرفاً به محبوبیت متکی نبودیم. ما میخواستیم مطمئن شویم که فهرست شامل عناوین آشنا و کتابهایی است که شرکتکنندگان ممکن است به تنهایی آنها را کشف نکرده باشند. این فهرست همچنین شامل انواع مختلفی از ژانرهای غیرداستانی گرفته تا رازآلود تا داستان های ادبی پیچیده تر بود.1
تیم تحقیقاتی این کتابها را از طریق آیپدهایی که در طول مدت مطالعه به امانت گذاشته بودند، بین شرکتکنندگان مسنتر در جامعه محلی توزیع کردند. آیپدها همچنین با یک برنامه سفارشی از قبل بارگذاری شده بودند که به شرکت کنندگان اجازه می داد پیشرفت خواندن خود را پیگیری کنند و به پرسشنامه های اضافی پاسخ دهند. شرکت کنندگان به مدت 90 دقیقه در روز، پنج روز در هفته، به مدت هشت هفته مطالعه کردند. یک گروه کنترل فعال جداگانه به جای خواندن، در حالی که پیشرفت آنها را با همان برنامه سفارشی ردیابی می کنند، پازل های کلمه را در iPad خود تکمیل کردند2.
استین-مورو گفت: «ما تا جایی که میتوانیم بین فعالیتها کنترل میکردیم، به جز «آب جادویی»، که در حال غرق شدن در یک داستان است.
در آغاز مطالعه، شرکتکنندگان به آزمایشگاه آموزش بزرگسالان در موسسه بکمن آمدند، جایی که از نظر مهارتهای شناختی مختلف، از جمله حافظه کاری و اپیزودیک، و همچنین سایر مهارتهای کلامی و خواندنی مورد ارزیابی قرار گرفتند. آنها در پایان هشت هفته دوباره بر روی این مهارت ها آزمایش شدند.
نتایج غیرقابل انکار بود: در مقایسه با گروه پازل، گروهی که به مدت هشت هفته کتاب می خواندند، بهبود قابل توجهی در حافظه فعال و حافظه اپیزودیک نشان دادند. به عبارت دیگر، این مطالعه نشان داد که خواندن منظم و متعهدانه مهارت های حافظه افراد مسن را تقویت می کند.
ارتباط علت و معلولی بین خواندن و حافظه چندین راه جدید را برای درمان های آینده برای بیماری هایی مانند بیماری آلزایمر باز می کند.
درگیر شدن کامل در کارهای محرکی که قبلاً در زندگی خود انجام می دهیم، نوید بیشتری وجود دارد. این احتمالاً بهترین راه برای حفظ توانایی ذهنی ما و خنثی کردن اثرات بیماری آلزایمر است.
کار آینده میتواند مزایای بلندمدت خواندن یا امکان تطبیق درمان خواندن با سلیقه شخصی افراد در کتابها را بررسی کند.
با این حال، در حال حاضر، پیام روشن است. چگونه با بالا رفتن سن از نظر ذهنی تیز بمانیم؟ کتاب بخوانید.
نتیجه گیری
در این مقاله، پژوهشگران از مؤسسه بکمن با همکاری کتابخانه عمومی شهر چامپین، از مزایای خواندن برای لذت بخشی در تقویت حافظه در بالغانها صحبت میکنند. پژوهشها نشان میدهند که خو1اندن منظم و مشغول به طور مداوم میتواند مهارتهای حافظه را تقویت کند و اساس برای رعایت بهتر مهارتهای روانی ما در سالهای پیری را فراهم کند.
- https://beckman.illinois.edu/about/news/article/2022/12/19/books-are-the-best-gift-for-your-brain-this -فصل تعطیلات ↩︎
- https://tall.education.illinois.edu/ ↩︎
همین امروز کلاس تندخوانی را شروع کنید
لیست کلاسهای درحال ثبت نام

دیدگاهتان را بنویسید